
Auteurs: Dr. Ellen Desie, Dr. Bary Muys & Dr. Wim Buysse (KU Leuven)
“Grote populaties van inheemse soorten die al lange tijd lokaal aanwezig zijn, beschikken vaak over een bredere genetische basis, wat hun aanpassingsvermogen vergroot.”
In het kader van klimaatslim bosbeheer is de keuze van boomsoorten en herkomsten voor bebossing en herbebossing een belangrijke uitdaging. Biedt het huidige soortenpalet en de genetische diversiteit van de lokale herkomsten voldoende ‘plasticiteit’ of aanpassingsvermogen om de klimaatverandering op te vangen? Of is het nodig om meer zuidelijke soorten en herkomsten te introduceren omdat ze misschien beter aangepast zijn aan onze toekomstige klimaatcondities?
Klimaatverandering
Klimaatverandering heeft nu al een grote impact op onze bossen en die zal alleen maar toenemen in de toekomst (Pörtner et al., 2022). Het is de afgelopen decennia significant warmer geworden (+0,2 °C per decennium wereldwijd) en deze gemeten opwarming gaat bijna dubbel zo snel in Europa (+0,38 °C per decennium in België) (KMI, 2020).
Klimaatmodellen voorspellen voor België een verdere toename van 2 °C tegen 2100 t.o.v. het huidige klimaatnormaal 1990-2020 (onder klimaatscenario SSP245, d.w.z. indien een klimaatbeleid geleidelijk wordt uitgevoerd), of zelfs tot 3,5 °C (klimaatscenario SSP585, business as usual, d.w.z. zonder bijkomende klimaatmaatregels) (KMI, 2020,2021). In beide gevallen komen verschillende inheemse boomsoorten onder druk te staan. De soorten die groeien hebben drie opties: uitsterven, zich aanpassen of zich verplaatsen naar gunstigere omstandigheden.

Foto 1: Een brede genetische basis vormt de essentie, en de beste garantie voor klimaatrobuustheid
“Klimaatmodellen voorspellen voor België een verdere toename van 2 C tegen 2100 t.o.v. het huidige klimaatnormaal 1990-2020."
Adaptatie van soorten
Het is niet eenvoudig om de klimaatgevoeligheid van boomsoorten te beoordelen. Gevoeligheid kan sterk afhangen van de context: zo zijn alleenstaande beuken en beuken in bosranden op zandgrond gevoelig voor droogte, maar blijken ze verrassend droogteresistent in bosverband op betere bodem. Daardoor kunnen publicaties mekaar sterk tegenspreken.
“Onderzoekers zijn het eens dat soorten- diversiteit en een grote genetische diversiteit binnen soorten cruciaal zijn voor een goede weerbaarheid van bossen.”
Door natuurlijke selectieprocessen zal steeds een (beperkt) deel van de nakomelingen beter aangepast zijn aan klimaatextremen die de moederbomen tijdens hun leven meemaken. Alle onderzoekers zijn het eens dat soortendiversiteit en een grote genetische diversiteit binnen soorten cruciaal zijn voor een goede weerbaarheid van bossen bij een veranderend klimaat.
Grote populaties van inheemse soorten die al lange tijd lokaal aanwezig zijn, hebben vaak een bredere genetische basis, wat het aanpassingsvermogen van deze soorten vergroot. Bovendien dragen deze inheemse soorten bij aan biodiversiteit door hun geassocieerde biodiversiteit, dit zijn soorten planten, schimmels, insecten, etc. die in nauwe samenhang leven met die boomsoort, en er vaak hun voortbestaan aan te danken hebben.
Autochtonie, dit is het voorkomen van populaties van inheemse soorten die zich na de ijstijden hebben gevestigd en die zich sindsdien natuurlijk hebben verjongd met lokaal genetisch materiaal, kàn een garantie zijn voor hogere resiliëntie, zoals het synchroon lopen van de bladuitloop met het uitkomen van vlindereitjes, en het voederen van rupsen door vogels aan hun jongen. Klimaatverandering kan deze relaties echter sterk verstoren.
Algemeen zijn autochtone populaties en lokale herkomsten van boomsoorten goed aangepast aan de historische klimatologische omstandigheden in Vlaanderen, waardoor ze plantmateriaal kunnen leveren dat minder vatbaar is voor vorst, ziekten en plagen. Voor soorten of herkomsten waarvan we vermoeden dat de genetische basis eerder beperkt is, kan men bij het aanplanten best verschillende aanbevolen herkomsten mengen. Bij populier wordt dit vertaald naar het werken met multiklonale aanplantingen.
Ook bij natuurlijke verjonging, waarbij er een vermoeden is dat de genetische basis van de ouderbomen beperkt is, kan je de genetische basis verbreden door actief bij te planten. De genetische connectiviteit in tijd en ruimte is vandaag de dag een belangrijk knelpunt.
“De genetische connectiviteit in tijd en ruimte is vandaag de dag een belangrijk knelpunt.”
Onze landschappen zijn sterk versnipperd waardoor de barrières voor spontane migratie enorm zijn toegenomen. Tegelijk zijn de ecologische opportuniteiten verminderd: de wildstand in Vlaanderen (vnl. ree, maar er beginnen zich ook populaties edelhert te vestigen) ligt enorm hoog waardoor werken met natuurlijke verjonging onder druk staat en er dus ook geen genetische adaptatie van inheemse soorten optreedt.
Behoud Vlaams bos voor klimaatadaptie in het Noorden
In tijden van klimaatverandering heeft het behouden van een brede genetische variatie in autochtone populaties ook een grote betekenis in een internationale context, van regio’s en metapopulaties, waarbij elke regio kan instaan voor het garanderen van de genetische variatie van meer noordelijke regio’s in de toekomst.
Migratie van soorten
In bergachtige gebieden zien we dat boomsoorten zich relatief eenvoudig kunnen aanpassen aan klimaatverandering door naar hogere hoogten te migreren. In vlakke gebieden zoals Vlaanderen daarentegen, is die aanpassing complexer. Hier moeten boomsoorten zich immers veel verder verplaatsen om dezelfde klimaatomstandigheden te vinden. Het blijft echter zeer waarschijnlijk dat verschillende soorten niet in staat zullen zijn zich tijdig aan te passen aan de veranderende omstandigheden en/of snel genoeg te migreren naar locaties met geschikte klimaatomstandigheden (Sittaro et al., 2017; Lenoir et al., 2020). Hoewel inheemse soorten klimaatverandering uit het verleden hebben doorstaan is het grote verschil de grote snelheid van de huidige klimaatverandering: de gemiddelde postglaciale noordwaartse migratie in Europa en Noord-Amerika lag in de grootte orde van 0,1 km/jaar (Pearson, 2006; Clark et al, 1998). De huidige klimaatsnelheid, de snelheid waarmee soorten hun leefgebied moeten verplaatsen om in gelijke klimaatomstandigheden te blijven naarmate het klimaat verandert, ligt voor loofbos en gemengd bos in de gematigde streken rond 0,35 km/jaar (Loarie et al 2009; Parks et al 2020). Dit gaat gemiddeld minstens drie keer sneller dan bossoorten in het West-Europese laagland zich kunnen aanpassen (zonder rekening te houden met habitatverlies, versnippering, hoge wildstand en andere milieudrukken). Daarom is het, naast het vrijwaren van genetische diversiteit, ook belangrijk te kijken hoe we migratie kunnen bevorderen, door de connectiviteit van onze systemen te verhogen en/of actief soorten te verplaatsen, om onze bosecosystemen zo veerkrachtig mogelijk te houden.
“Geassisteerde migratie kan een hulpmiddel zijn voor klimaatadaptatie van multifunctionele bossen, en zelfs als een laatste redmiddel gezien worden in het natuurbeheer.”
Geassisteerde migratie is van alle tijden. Sinds de prehistorie heeft de mens bepaalde boomsoorten verplaatst binnen en over continenten. In de 19de eeuw werden enkele snelgroeiende exoten voor productiebosbouw ingevoerd. Geassisteerde migratie kan een hulpmiddel zijn voor klimaatadaptatie van multifunctionele bossen (Messier et al., 2019), en zelfs als een laatste redmiddel gezien worden in het natuurbeheer (Prober et al., 2019). We kunnen bepaalde uitheemse soorten en herkomsten inzetten in onze bosecosystemen in Vlaanderen (Messier et al., 2015).
“In bergachtige gebieden zien we dat boomsoorten zich relatief eenvoudig kunnen aanpassen aan klimaatverandering door naar hogere hoogten te migreren.”
Discussie over inheems of niet?
“Hoe dan ook houden boomsoorten zich niet aan staatsgrenzen.”
Volgens de ecologische definitie is een soort inheems als ze voorkomt in haar natuurlijk verspreidingsgebied (den Ouden et al., 2010). Het natuurlijk verspreidingsgebied is het geografische gebied waarin een soort oorspronkelijk voorkomt en zich van nature heeft verspreid zonder directe menselijke invloed. Soms wordt daar nog een termijn bij geplaatst, bijvoorbeeld minstens 1.500 jaar (Kornas, 1983), 5.000 jaar (Park, 2007) of sinds de laatste ijstijd.
In het Bosdecreet wordt een inheemse boom- of struiksoort gedefinieerd als een boom- of struiksoort, die van nature voorkomt in een bepaalde streek of regio. Een lijst van bomen en struiken die juridisch als Vlaams inheems beschouwd worden is te vinden in bijlage 1 van het Besluit van de Vlaamse Regering van 2 oktober 2015 betreffende het verlenen van subsidies voor bebossing en voor herbebossing (B.S. 20/11/2015). Deze lijst is door experten opgesteld maar over een aantal soorten blijft er wetenschappelijke discussie. Want de exacte definitie is nooit echt scherp en duidelijk afgelijnd geweest en vaak gebaseerd op arbitraire grenzen in tijd en ruimte (Hermy, 2020; Pereyra, 2020). Zo komt het dat wilde appel in Vlaanderen inheems beschouwd wordt en tamme kastanje niet, terwijl ze beiden hier al lang actief door mensen zijn verspreid als voedselplant. Noorse esdoorn wordt beschouwd als inheems in België maar niet van nature voorkomend in Vlaanderen hoewel hij in de grenszone tussen de regio's mogelijks wel voorkwam.
Hoe dan ook houden boomsoorten zich niet aan staatsgrenzen, het ‘natuurlijk’ verspreidingsgebied van een soort kan dus uitbreiden, inkrimpen, verschuiven, fragmenteren of desintegreren tot de soort enkel in een aantal refugia overblijft (Hobbs et al. 2018). In onze studie waarschuwen de onderzoekers dat het begrip "Vlaams inheems" te rigide is in een tijd waar het klimaat zo snel verandert dat ook soortenniches mee opschuiven en wordt voorgesteld om het over "inheems in het west-palearctisch gebied" of kortweg "Europees inheems" te hebben. Dit ruimere begrip omvat een bredere soortenpool, waarbij niettemin in grote mate sprake is geweest van co-evolutie tussen deze boomsoorten onderling en met hun geassocieerde biodiversiteit.
De aanpassingen in wetgeving en beleid waren gedurende meer dan 30 jaar vooral tegen monoculturen met niet inheemse soorten gericht. Maar vandaag wordt een soort die buiten zijn natuurlijk verspreidingsgebied wordt gevonden bijna automatisch als niet-inheems en dus potentieel invasief of schadelijk beschouwd. Dit terwijl verspreidingsgebieden volop aan het verschuiven zijn. Daarom pleit een groeiend aantal wetenschappers voor een aanpak waarbij men de boomsoortensamenstelling van stabiele natuurlijke en half natuurlijke ecosystemen in een goede staat probeert te behouden, maar daarnaast te aanvaarden dat dit in veel gevallen niet zal lukken. In die gevallen moeten we ons voorbereiden om ze via soortselectie en beheer in de richting van nieuw-samengestelde bosecosystemen te leiden die het bosmicroklimaat nodig voor bosbewonende organismen bewaren en het leveren van bosecosysteemdiensten blijven garanderen (Davis et al., 2011; Jackson & Hobbs, 2009).
Risico’s bij geassisteerde migratie
Geassisteerde migratie is echter niet zonder risico: de mogelijke invasiviteit en groeiplaatsgeschiktheid van soorten moet door voorafgaande selectie en verdere monitoring nauwkeurig worden geëvalueerd. Hierna volgen de belangrijkste argumenten tegen geassisteerde migratie.
Invasiviteit
Risico's voor ecosystemen ten gevolge van geassisteerde migratie bestaan uit mogelijke invasiviteit, hybridisatie, competitie met inheemse soorten en onvoorziene veranderingen in het milieu. Gepaste beheermaatregelen, zoals het vermijden van monoculturen en het controleren van ziekteverspreiding, zijn van essentieel belang. Het is noodzakelijk om de verschillende eigenschappen van boomsoorten in rekening te nemen, waaronder hun potentieel voor invasiviteit. In een tijd van verschuivende ecologische niches, is het cruciaal om nieuwe soorten niet automatisch als invasieve exoten te bestempelen.
Heldere definities voor "inheems" en "invasief", los van elkaar, ontbreken momenteel. Hoewel invasiviteit per definitie wordt geassocieerd met exotische soorten, vertonen inheemse boomsoorten met een pionier-karakter ook vaak een expansief en dominant gedrag (men spreekt dan van een “native invader” sensu Carey et al. 2012 ): inheemse Grove den en Ruwe berk kunnen bijvoorbeeld problematisch zijn in heidevegetaties (Nyssen et al. 2023). Het is van belang om definities correct te hanteren in functie van de vraagstelling maar uiteindelijk moet de beheerder beslissingen maken over soortenbeheer in functie van (huidige/toekomstige) impact, wat belangrijker is dan de 'afkomst' van een soort. Om de mogelijke impact te evalueren kan men gebruik maken van risicoanalyseprotocollen.

Foto 2: De nacht van 22 april 2024 werd in Kleine Brogel -1,5 °C onder thermometerhut gemeten. Aan de grond daalde de temperatuur tussen -3 en -5 °C. Onbeschutte jonge eikjes en beukjes (lokaal aanbevolen herkomsten) leden schade, maar niet onder scherm.
Geassocieerde biodiversiteit
Voor veel nieuwe boomsoorten blijft geassocieerde biodiversiteit een kennislacune terwijl we voor lokaal inheemse boomsoorten een betere kennis hebben (zeer hoog voor bijvoorbeeld zomereik, erg laag voor bijvoorbeeld taxus). In principe gaan we er van uit dat hoe langer een boomsoort in een bepaald gebied voorkomt, hoe hoger het aantal ermee geassocieerde soorten in een bepaald habitat ("alpha-diversiteit"). Hieruit kunnen we niet zomaar besluiten dat de geassocieerde diversiteit op inheemse soorten steeds hoger is dan op exoten. Ook blijkt dat er relatief snelle aanpassingen optreden, waarvan sommige evolutionair verankerd zijn. Het aantal bladetende insecten op Amerikaanse vogelkers is de afgelopen 150 jaar verdubbeld tot 69 soorten terwijl dit aantal op Europese vogelkers lager ligt (39 soorten) en stabiel bleef (Schilthuizen et al., 2016). Dit werd ook vastgesteld bij een reeks andere ingeburgerde boomsoorten (Nyssen, 2023). De meest bepalende factor is echter de abundantie, of er veel exemplaren van een boomsoort in een bepaald gebied voorkomen. In de nabije toekomst wordt het met behulp van eDNA analyse ("environmental DNA") mogelijk om de totale soortendiversiteit op boomsoorten te bepalen, dus niet enkel insecten. En dan komen er hoogstwaarschijnlijk nog meer nuanceringen aan het licht.
Late vorst
Zuiderse soorten kunnen minder bestand zijn tegen nachtvorst. Het aantal vorstdagen in Vlaanderen vermindert maar (nacht)vorst blijft in principe mogelijk tot ongeveer de IJsheiligen (11 tot 14 mei). Het uitlopen van knoppen wordt bij soorten uit gematigde streken over het algemeen bepaald door twee variabelen: temperatuur en daglengte. Bij korte daglengtes is er meer warmte nodig om uit te lopen, dit beschermt de boom tegen te vroeg uitlopen. Bij lange daglengtes is er minder warmte nodig, dit beschermt de boom tegen te laat uitlopen. Voorafgaand is bij de meeste soorten ook een bepaald minimum aantal koude dagen nodig. Zowel het moment van uitlopen als het tijdstip van de laatste voorjaarsvorst valt steeds vroeger, maar uit een modellering in Wallonië van beuk, wintereik en es en Europese lork blijkt dat er tot nu toe een veilige marge blijft tussen laatste kans op vorst en eerste kans op uitlopen (Borlon et al., 2022). Bovendien is het vooral de exacte locatie van de bomen die bepaalt of vorstschade optreedt: een laaggelegen kom in het landschap, droge zandgrond, kaalvlakte bij windstil weer en heldere hemel, ... ongeacht of dit lokale of zuidelijke herkomsten zijn. Dit heeft dus in eerste instantie met de manier van bosaanleg of herbebossing te maken: plant onder scherm of zorg voor een snelle kroonsluiting door voldoende dicht te planten.
Zijn de risico’s groter dan het risico van niets doen?
We moeten ons echter ook de vraag stellen wat het risico is van migratie tegenhouden (bv. het bestrijden van natuurlijke vestiging van Steeneik, Quercus ilex, in bossen aan de kust, buiten waardevolle duinvegetaties waar controle van deze en andere boomsoorten nodig is) en/of het niet assisteren van boomsoorten in soortenarme en klimaatgevoelige bossen.
Foto 3 toont op de voorgrond een nieuwe aanplant in open vlakte na massale bossterfte in het Hardtwald in de Rijnslenk van Baden Württemberg. De uitgestrekte dennenmengbossen op zandgrond zijn vergelijkbaar qua soortensamenstelling met onze Kempen, doch 400 km zuidelijker, en geven een idee hoe klimaatverandering er over een paar decennia kan uitzien in Vlaanderen. Huidige toestand: berk verdwijnt volledig, grove den en douglas sterven massaal, van beuk sterven de toppen af maar hij houdt voorlopig nog stand, eik verliest vitaliteit; Amerikaanse eik, Amerikaanse vogelkers en Robinia staan in goede gezondheid, hemelboom profiteert in sterfteplekken.

Foto 3: Nieuwe aanplant in open vlakte na massale bossterfte in het Hardtwald in de Rijnslenk van Baden-Württemberg.
Risico’s bij geassisteerde migratie
Bij geassisteerde migratie worden boomsoorten verplaatst binnen hun verspreidingsgebied of zelfs daarbuiten, om hen een betere kans op overleving te geven in veranderende klimaatomstandigheden. Om te experimenteren zijn een aantal zaken belangrijk. Zo zal nooit een enkele soort dé oplossing bieden, wel soortengroepen en gemeenschappen. We moeten hierbij streven naar soortendiversiteit, functionele diversiteit en functionele redundantie binnen het bos: we moeten op zoek gaan naar soortencombinaties die elkaar aanvullen in hun functies.
Vroeger selecteerde men boomsoortenkeuzes op basis van ervaring en intuïtie. De veranderende context van klimaat maken dat intuïtie en ervaring moeilijk nog kunnen bevatten wat goed zou werken.
Vandaag beschikken we over soortenverspreidingsmodellen die advies geven over welke bomen te selecteren.
In het kader van die uitdaging werd KU Leuven in opdracht van ANB gevraagd een wetenschappelijk onderbouwde lijst op te maken van boomsoorten die voor ons toekomstig klimaat geschikt zijn, ook het Klimaatbomenproject genoemd.
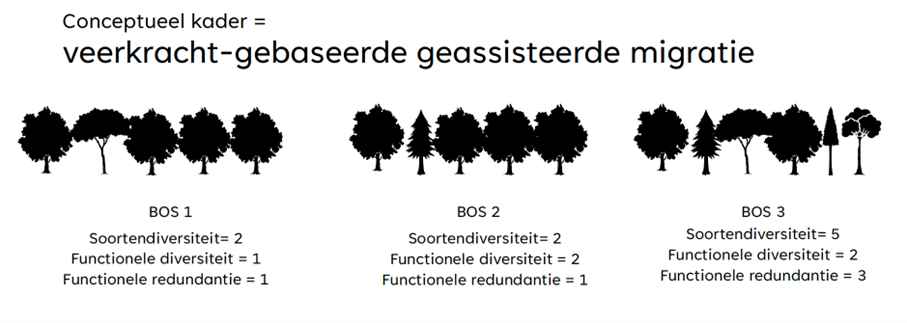
Figuur 1: Veerkracht-gebaseerde geassisteerde migratie